





Así pues, la otra vertiente de la actividad cerebral (eléctrica) hasta hoy en comparación ha sido poco estudiada como posible método de intervención regulador o potenciador de la actividad cerebral, sin embargo si ha sido estudiado como método de diagnóstico neurológico a través de los electroencefalogramas (EEG) ya desde hace algunos años cuando H. Berger logro medir por primera vez la actividad cerebral identificando las frecuencias alfa y las beta como se citó en Compston (2010).
Las neuronas comparten las mismas características y tienen las mismas partes que otras células en el cuerpo, pero pueden transmitir mensajes entre sí de largas distancias a través de sus procesos axonales. Al igual que prácticamente cualquier célula en el cuerpo, las neuronas tienen una alta concentración de iones de potasio (K) y cloruro (Cl) en el interior así como iones de sodio (Na) 2+ y calcio (Ca) en el exterior. Esta disposición produce una pequeña batería que mantiene una diferencia de tensión de -60 milivoltios con respecto al mundo exterior de la membrana celular.
Las neuronas se comunican entre sí mediante el envío de señales electroquímicas de la terminal pre-sináptica del axón de una célula a la terminal post-sináptica de las dendritas de otras células.
Las sustancias químicas emitidas por las uniones sinápticas pueden excitar (de-polarizar) o inhibir (hiper-polarizar) la membrana neuronal. La excitación de la membrana neural cambia su permeabilidad a los iones en el líquido extracelular. Típicamente, una célula en reposo es altamente permeable a los iones de potasio y relativamente impermeable a los iones sodio. La concentración de iones de potasio en el interior de la célula es 35 veces mayor que la concentración de iones de potasio fuera de la célula. Este gradiente de concentración tiende a conducir los iones de potasio fuera de la célula.
La difusión hacia el exterior resultante de iones de potasio, que son positivamente cargados, conduce a un exceso de iones de carga negativa dentro de la célula. El desequilibrio resultante de la carga a través de la membrana celular impide la salida de más iones de potasio. Los iones de sodio se alinean en el exterior de la membrana celular, pero la membrana no es suficientemente permeable para neutralizar la carga interior negativa resultante del flujo de iones de potasio.
La membrana al continuar actuando como una barrera que separa las cargas en cada uno de sus lados, a su vez, se polariza, así neurotransmisores excitatorios, tales como la acetilcolina, se adhieren a un sitio receptor en la dendrita en la membrana post-sináptica, resultando en un aumento de la permeabilidad de la membrana celular a los iones de sodio.
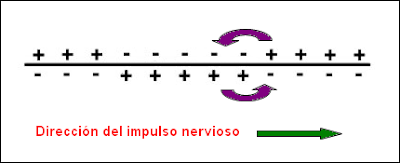
La afluencia resultante de carga positiva establece un potencial exitatorio post-sináptico (PEP). Los PEP son pequeños, por lo general solo llegan a los 5 mV, lo cual no es suficiente para desencadenar un potencial de acción, o sea, el cambio necesario en el potencial de la membrana para enviar una señal a lo largo de un axón a la terminal sináptica causando la liberación de neurotransmisores en el hueco post-sináptico. Mas como se mencionó anteriormente, la mayoría de las neuronas son multipolares, y por lo tanto tienen muchas dendritas cuyas cargas crean una suma de PEP suficientes para desencadenar un potencial de acción (aproximadamente 20-30 mV) (Figura 1.3).
Figura 1.3
Fuente: http://1.bp.blogspot.com/_TYKXEPKoytc/S8MjPOhDS1I/AAAAAAAADFk/_RaXlS6xRik/s400/N7.PNG
{kind=link}
Estas sumas de cargas pueden ser espaciales o temporales. La sumatoria espacial es la suma de varios PEP que se producen simultáneamente en diferentes sitios. La sumatoria temporal es la suma que se produce como resultado de sucesivos cambios potenciales en un solo sitio de tal manera que varios PEP se superponen unos sobre otros.
Los potenciales de acción constituyen las señales por el cual el cerebro recibe, analiza y transmite información. Estas señales son altamente estereotipadas en todo el sistema nervioso, a pesar de que pueden ser iniciadas por una gran variedad de eventos del medio ambiente, desde la luz hasta el contacto mecánico, y desde olores a ondas de presión.
Así, las señales que transmiten información acerca de la visión son idénticos a los que llevan la información acerca de los olores. Aquí encontramos un principio fundamental de la función cerebral. La información transmitida por un potencial de acción no está determinada por la forma de la señal, sino por la vía en que la señal viaja en el cerebro. El cerebro analiza e interpreta los patrones de señales eléctricas entrantes y de esta manera crea nuestras sensaciones cotidianas. (Kandel, 2000).
La importancia de estas frecuencias cerebrales cada día se hace más evidente, Buzsáki (2012) nos dice que “La perpetua actividad de la corteza cerebral está sustentada en gran medida por la variedad de oscilaciones que genera el cerebro, las que abarcan un número de frecuencias y sitios anatómicos, así como de correlatos conductuales.”
En el mismo sentido, Nunes y Srinivasan (2006) anteriormente habían comentado que “los potenciales eléctricos o electroencefalogramas (EEG) leídos en el cuero cabelludo proporcionan estimaciones de acción sináptica a gran escala estrechamente relacionados con el comportamiento y la cognición. Por lo tanto, el EEG puede proporcionar una genuina ventana hacia la mente”
Erol Basar (1999) afirmaba que “las oscilaciones del EEG permiten el análisis de las funciones cognitivas y sensoriales en su conjunto”
Algunos autores proponen que los ritmos o frecuencias cerebrales implican una especie de sintaxis neural (Buzsaki y Watson, 2012)
Hoy reconocemos 5 bandas de frecuencias que estas están relacionadas con diferentes procesos cognitivos (tabla 1) hay quienes hace divisiones adicionales como la frecuencia Mu que cae dentro de la alfa (Steriade, Gloor y Llinas, 1990)